Für uns Menschen ist das Sehen der wichtigste Sinn. Unsere Augen sind hochempfindlich, aber gleichzeitig enorm anpassungsfähig. Die Erfindung des Sehens brachte in der Evolution gewaltige Vorteile. Wer sehen kann, vermag Nahrung, Gefahr und mögliche Partner aus grosser Distanz zu erkennen, was wiederum neue Wege zur Kommunikation und zur Orientierung eröffnet.
Lernziele
- Sie können den Prozess der Akkommodation erläutern.
- Sie kennen den Aufbau der Retina.
- Sie kennen die Eigenschaften der Photorezeptoren und ihre Verteilung in der Retina.
- Sie können die Phototransduktion in den Photorezeptoren erklären.
- Sie verstehen wie es durch laterale Hemmung zur Kontrastverschärfung kommt.
9.1 – Licht
Licht ist elektromagnetische Energie, die in Form von Wellen emittiert wird. Nur ein winziger Ausschnitt des elektromagnetischen Spektrums ist für uns als Licht sichtbar. Die kürzeste sichtbare Wellenlänge von ca. 400 nm erscheint uns blau-violett, die längste noch sichtbare von ca. 700 nm rot-braun. Die Wellenlängen dazwischen entsprechen den anderen Farben des Regenbogens. Im Spektrum elektromagnetischer Strahlung wird das sichtbare Licht von der längerwelligen infraroten und der kürzerwelligen ultravioletten Strahlung flankiert. Ultraviolettes Licht hat bereits genügend Energie, dass es die Hornhaut des Auges in Form eines “Sonnenbrands im Auge” schädigen kann. Die von der Sonne emittierte Mischung der Wellenlängen erscheint dem Menschen als weisses Licht (polychromatisches Licht), während Licht einer einzigen Wellenlänge (monochromatisches Licht) als einzelne Farbe wahrgenommen wird.
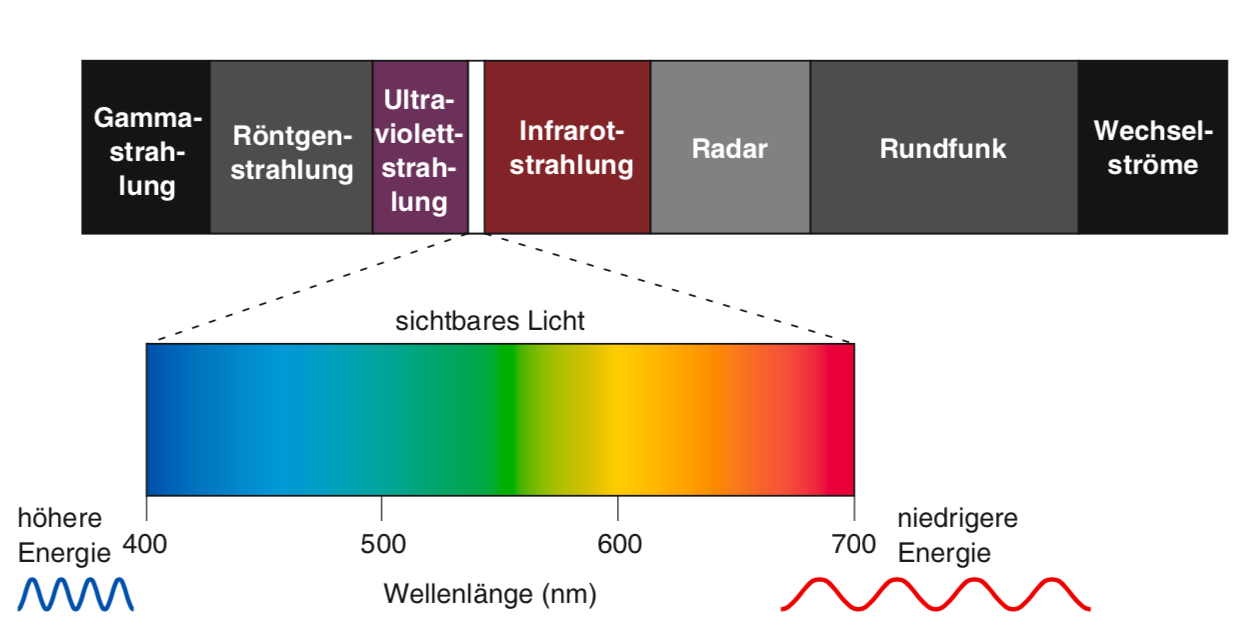
Abbildung 9.1 – Das elektromagnetische Spektrum. Nur elektromagnetische Strahlung mit einer Wellenlänge zwischen 400 und 700 nm ist für das menschliche Auge sichtbar. Innerhalb dieses sichtbaren Spektrums erscheinen die unterschiedlichen Wellenlängen als unterschiedliche Farben. Quelle: Mark Bear et al.: Neurowissenschaften, 4.Auflage, Springer Spektrum, 2018, S. 311
Reflexion ist das Zurückwerfen der Lichtstrahlen von einer Oberfläche. Die Art, wie ein Lichtstrahl reflektiert wird, hängt von dem Winkel ab, in dem er auf die Oberfläche trifft. Absorption ist die Übertragung von Lichtenergie auf einen Partikel oder eine Oberfläche. Flächen, die schwarz erscheinen, absorbieren die Energie aller sichtbaren Wellenlängen. Einige Verbindungen absorbieren die Lichtenergie nur in einem begrenzten Wellenlängenbereich und reflektieren die übrigen Wellenlängen. Diese Eigenschaft ist die Basis für die Farbpigmente in Farben.
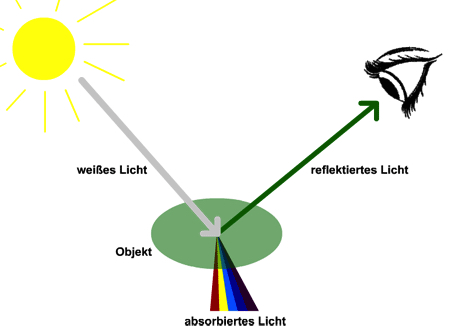
Abbildung 9.2 – Eine grüne Oberfläche absorbiert Licht aller Wellenlänge. Ausgenommen ist das grünes Licht, welches reflektiert wird. Quelle: https://www.univie.ac.at/mikroskopie/1_grundlagen/faerbung/einleitung.htm, 10.12.2018
Die Unterscheidung von Objekten beruht beim Sehen vor allem auf der Wahrnehmung von Farbkontrasten (unterschiedliche Wellenlängen), die durch unterschiedliche Reflexion oder Aussendung von Licht durch Objekte entstehen. Weiter müssen die Objekte, damit man sie vom Hintergrund unterscheiden kann, einen Hell-Dunkel-Kontrast aufweisen. Er wird durch die Leuchtdichte (Lichtstrom pro Fläche), die von einem Objekt ausgeht, bestimmt.
9.2 – Das Auge
Anatomie des Auges
Sie haben die Anatomie des Auges bereits in der Vorlesung von L.Slomianka kennengelernt. Beschriften Sie zur Wiederholung die wichtigsten anatomischen Strukturen.
9.3 – Bildentstehung im Auge
Das Auge sammelt Lichtstrahlen, die von Objekten in der Umgebung ausgestrahlt oder reflektiert werden, und fokussiert sie auf der Retina (=Netzhaut), um Bilder zu erzeugen. Um Objekte in den Fokus zu bringen, wird die vereinte Brechkraft von Cornea (=Hornhaut) und Linse benötigt. Wie in Abb. 9.3 dargestellt, gelangt Licht, das senkrecht auf die Oberfläche der Cornea trifft, auf geradem Weg auf die Retina, aber Lichtstrahlen, die nicht rechtwinklig auf die gekrümmte Oberfläche der Cornea treffen, werden so gebrochen, dass sie auf der Retina am Augenhintergrund zusammenlaufen. Die Strecke zwischen der Brechungsoberfläche und dem Punkt, an dem die Lichtstrahlen zusammentreffen (der sogenannte Brennpunkt), wird als Brennweite bezeichnet. Sie ist abhängig von der Krümmung der Hornhaut – je stärker die Krümmung, desto kürzer ist die Brennweite.

Abbildung 9.3 – Brechung durch die Cornea (=Hornhaut). Die Cornea muss eine ausreichende Brechkraft besitzen, um Licht auf der Netzhaut am Augenhintergrund zu fokussieren. Quelle: Mark Bear et al.: Neurowissenschaften, 4.Auflage, Springer Spektrum, 2018, S. 317
9.4 – Akkommodation
Obwohl die Cornea den grössten Teil der Lichtbrechung bewirkt, spielt die Linse eine wichtigere Rolle bei der Erzeugung von scharfen Abbildern von Gegenständen, die weniger als ~9m vom Auge entfernt sind. Bei näheren Objekten wird eine grössere Brechkraft benötigt, um sie auf der Retina zu bündeln. Diese zusätzliche Brechkraft wird durch die Veränderung der Linsenform erzielt, ein Vorgang, der Akkommodation genannt wird (Abbildung 9.5).
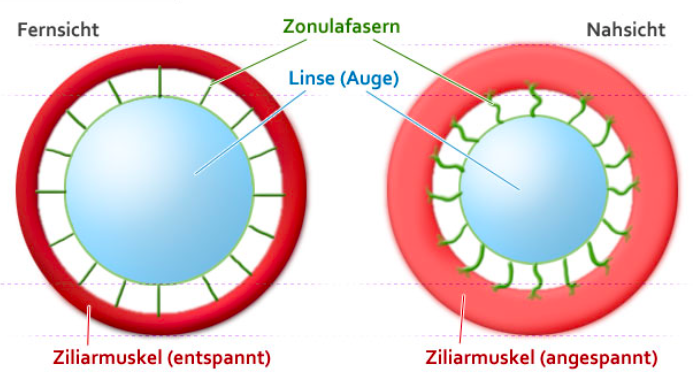
Abbildung 9.4 – Ziliarmuskel Quelle: lasikon.de
Der Ziliarmuskel bildet einen Ring in den die elastische Linse über Aufhängebänder (Zonulafasern) eingespannt ist. Ist der Zilliarmuskel entspannt, stehen die Aufhängebänder unter Spannung, die Linse wird auseinadergezogen und dadurch abgeflacht. Während der Akkommodation kontrahiert sich der Ziliarmuskel und schwillt an. Dadurch verkleinert sich die Fläche innerhalb des Muskelrings, und die Aufhängebänder entspannen. Als Folge rundet sich die Linse aufgrund ihrer natürlichen Elastizität ab. Diese Rundung verstärkt die Wölbung der Linsenoberfläche und vergrössert auf diese Weise die Brechkraft.
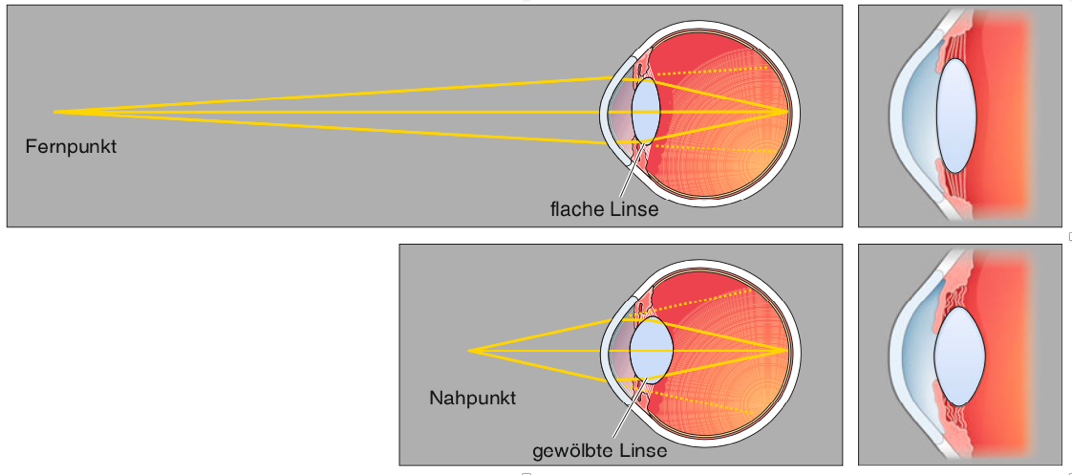
Abbildung 9.5 – Akkommodation durch die Linse. Zur Fokussierung eines weit entfernten Punktes wird eine relativ geringe Brechung benötigt. Der Ziliarmuskel entspannt sich, die Zonulafasern werden gedehnt, und die Linse wird abgeflacht. Nahe Objekte erfordern eine grössere Brechung. Dazu kontrahiert der Ziliarmuskel, sodass weniger Zug auf die Zonulafasern wirkt. Quelle: Mark Bear et al.: Neurowissenschaften, 4.Auflage, Springer Spektrum, 2018, S. 318
Die Fähigkeit zur Akkommodation verändert sich im Laufe des Lebens. Kinder können mit ihren Augen Objekte direkt vor ihrer Nase fokussieren, während viele Menschen mittleren Alters Dinge, die eine Armeslänge entfernt sind, schon nicht mehr scharf sehen können. Alterssichtigkeit beruht auf einer Verhärtung der Linse und kann damit erklärt werden, dass zwar während des gesamten Lebens neue Linsenzellen produziert, aber keine abgebaut werden. Die verhärtete Linse ist weniger elastisch, sodass sie ihre Form kaum noch ändern und sich daher nicht ausreichend wölben kann umd nahe Objekte scharf auf der Netzhaut abzubilden.
9.4.1 – Kurzsichtigkeit und Weitsichtigkeit
Bei Myopie handelt es sich um die sogenannte Kurzsichtigkeit. Sie zeichnet sich durch einen zu langen Augapfel (=Bulbus) aus. So liegt der Brennpunkt vor der Retina und das Bild auf der Retina ist unscharf. Durch ein konkaves bzw. zerstreuendes Glas oder Linse kann wieder eine normale Fernsicht hergestellt werden. Häufig wird Kurzsichtigkeit durch sehr kurze Lese- und Schreibdistanzen induziert (<30cm). Dabei sind vorallem Kinder und Jugendliche betroffen, da häufiges Lesen und Schreiben in dieser Distanz das Wachstum des Augapfels fördert. Bei der Hypermetropie, auch Weitsichtigkeit genannt, liegt die scharfe Bildebene hinter der Fovea, weil der Augapfel im Verhältnis zur Brechkraft zu kurz ist. Korrigiert wird dies mit einem Sammelglas.
9.5 – Retina
Mit einem Augenspiegel kann ein Augenarzt auf den Augenhintergrund mit der Retina blicken und dort mehrere Strukturen erkennen. Am sogenannten blinden Fleck, der Papille, treten Blutgefässe in die Retina ein und Fasern des Sehnervs aus der Retina aus. Hier liegen keine Lichtsinneszellen, weshalb wir an dieser Stelle blind sind. Am gelben Fleck befindet sich die Stelle des schärfsten Sehens, die sogennate Fovea.
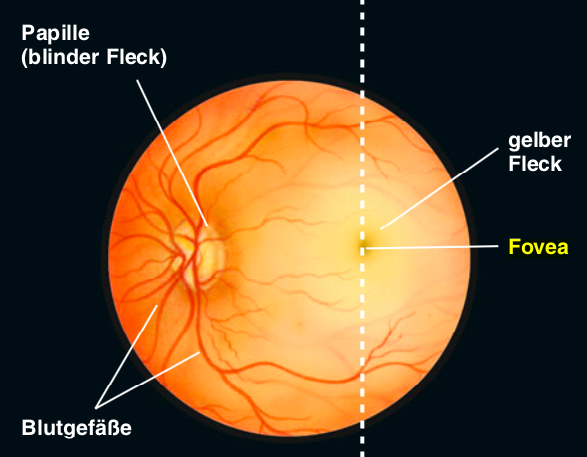
Abbildung 9.6 – Die Netzhaut, betrachtet durch einen Augenspiegel. Quelle: Mark Bear et al.: Neurowissenschaften, 4.Auflage, Springer Spektrum, 2018, S. 313
9.5.1 – Fovea centralis
Die Retina ist eine Ausstülpung des Gehirns, quasi ein vorgeschobener Posten. Sie enthält nicht nur die Photorezeptoren, sondern auch ein direkt dahinter geschaltetes neuronales Netzwerk. Hier wird die Lichtinformation im Sinnesorgan bereits intensiv verarbeitet, bevor sie an das Gehirn weitergeleitet wird. Die Funktionsweise dieser neuronalen Zellen lernen sie im Kapitel 9.7.1 „Neuronale Verschaltung und laterale Hemmung“ im Detail kennen. Erstaunlicherweise liegen die Photorezeptoren auf der Seite der Retina, die dem Licht abgewandt ist. Bevor das Licht die Photopigmente in den Photorezeptoren erreicht, muss es also zuerst die anderen Retinaschichten durchdringen. Am gelben Fleck, der Stelle des schärfsten Sehens, wurden die neuronalen Zellen der Retina einfach zur Seite geschoben und bilden eine Sehgrube. Der Punkt des schärfsten Sehens ist genau in der Mitte der Sehgrube und wird Fovea centralis genannt.

Abbildung 9.7 – Fovea. Quelle: Mark Bear et al.: Neurowissenschaften, 4.Auflage, Springer Spektrum, 2018, S.326
Diese strukturelle Spezialisierung sorgt für eine maximale Sehschärfe in der Sehgrube, weil alle anderen Zellen, die das Licht streuen und das Bild unscharf aussehen lassen könnten, aus dem Weg geräumt sind. Photorezeptoren der Fovea centralis leiten ihre Informationen häufig nur an eine nachgeschaltete neuronale Zelle weiter. Diese Zellen besitzen daher kleine rezeptive Felder, die eine hohen Auflösung ermöglichen. In dem gesamten restliche Teil der Retina konvergiert die Information von relativ vielen Photorezeptoren auf eine nachgeschaltete Zelle. Deshalb besitzen diese Zellen grosse rezeptive Felder und eine niedrigere Auflösung. Warum fällt uns im täglichen Leben nicht auf, dass unsere Retina nur im Zentrum ein scharfes Bild vermittelt? Ganz einfach: weil wir unsere Augen ständig bewegen. Sobald ein Objekt im peripheren Gesichtsfeld auftaucht, wird es zuerst unscharf wahrgenommen und so an das Gehirn gemeldet. Das Gehirn löst sofort eine Reihe von Augenbewegung aus wodruch das Objekt Stück für Stück „gescannt“ und scharf auf der Fovea centralis abgebildet wird. Diese Information wird analysiert und von unserem Gehirn so zusammengesetzt, dass das Objekt als eine Einheit wahrgenommen wird. Der Grund für diesen Aufbau der Retina ist eine Reduktion der Information. Wenn wir mit der gesamten Retina scharf sehen würden, könnte unser Gehirn die Informationsflut, die von so vielen Bildpunkten kommt, gar nicht bewältigen.
9.6 – Photorezeptoren
In den Photorezeptoren auf der Rückseite der Retina findet die Umwandlung von elektromagnetischer Strahlung in neuronale Signale statt. Jeder Photorezeptor besteht aus vier Regionen: einem Aussensegment (dem Augenhintergrund zugewandte Seite), einem Innensegment (dem Lichteinfall zugewandte Seite), einem Zellkörper und einer synaptischen Endigung. Das Aussensegment enthält einen Stapel von Membranscheibchen. Lichtempfindliche Photopigmente in diesen Scheibchen absorbieren Licht und lösen dabei eine Veränderung im Membranpotenzial des Photorezeptors aus. Es gibt zwei Arten von Photorezeptoren in der Retina. Man kann sie leicht anhand der Erscheinungsform ihrer Aussensegmente unterscheiden: Stäbchen haben ein langes zylindrisches Aussensegment mit vielen Membranscheibchen. Zapfen besitzen ein kürzeres, sich zuspitzendes Aussensegment mit weniger Membranscheibchen. Die grössere Anzahl Membranscheibchen in den Stäbchen machen diese über 1000-mal lichtempfindlicher als die Zapfen.
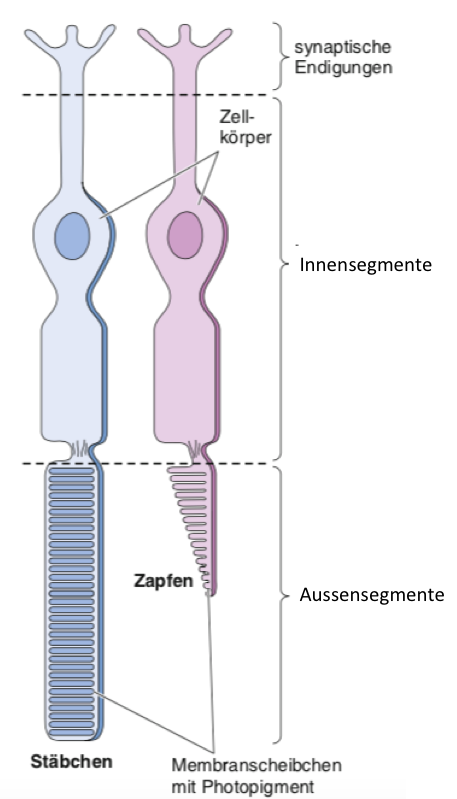
Abbildung 9.8 – Stäbchen und Zapfen. Quelle: Mark Bear et al.: Neurowissenschaften, 4.Auflage, Springer Spektrum, 2018, S. 324
Die Stäbchen sind so empfindlich, dass sie auf einzelne Lichtquanten reagieren. Mit ihnen sehen wir bei Sternenlicht. Dann empfängt jedes Stäbchen in unserem Auge lediglich ein oder wenige Lichtquanten pro Stunde. Wenn bei Mondlicht mehr Quanten vorhanden sind als bei Sternenlicht, verstärkt das Stäbchen das Signal weniger stark als bei schwachem Licht und passt sich so den Gegebenheiten an. Trotz dieser Anpassungsfähigkeit stossen die Stäbchen bei hellerem Licht an ihre Grenzen. Ihre Antwort wird ab einer bestimmten Reizintensität nicht mehr grösser – sie ist gesättigt. Die Stäbchen sind bei diesen Gegebenheiten zum Sehen ungeeignet. Wären wir bei hellem Licht weiterhin ausschliesslich auf die Stäbchen angewiesen, würden wir das Gefühl haben, ständig geblendet zu werden. Glücklicherweise kommen nun die Zapfen ins Spiel. Sie sind von Anfang an weniger empfindlich und können ihre Empfindlichkeit zudem über einen grossen Belichtungsbereich hinweg anpassen. Die Zapfen decken damit den ganzen Bereich des Tagessehens ab. Während alle Stäbchen das gleiche Photopigment enthalten, gibt es drei Arten von Zapfen, jede mit einem anderen Pigment. Die Unterschiede zwischen den Pigmenten führen dazu, dass die drei Arten von Zapfen unterschiedliche spektrale Empfindlichkeiten besitzen und damit auf unterschiedliche Farben reagieren, d.h. die Zapfen sind für die Farbwahrnehmung verantwortlich.
Der Aufbau der Retina variiert von der Fovea centralis bis zur Retinaperipherie. Die meisten der rund fünf Millionen Zapfen befinden sich in der Fovea, also dem Bereich der Retina, der ein scharfes Bild vermittelt. Zur Peripherie hin verringert sich die Anzahl an Zapfen beträchtlich. Während in der Fovea gar keine Stäbchen vorhanden sind, enthält die periphere Retina sehr viel mehr Stäbchen als Zapfen.
Welche Konsequenzen hat die Verteilung der Stäbchen und Zapfen auf der Retina?
9.7 – Phototransduktion
Photorezeptoren unterscheiden sich funktionell von den Sensoren, die wir bereits bei anderen Sinnesorganen kennengelernt haben. Während alle bisherigen Sensoren durch einen Reiz depolarisiert werden, ist das bei den Photorezeptoren genau anders herum: Photorezeptoren sind im Dunkeln depolarisiert und reagieren auf einen Lichtreiz mit Hyperpolarisation. Dieser Vorgang der Photortransduktion wird vor allem durch postive geladene Natrium-Ionen (Na+) gesteuert. Die Zellmembran des Photorezeptors enthält viele Na+ Kanäle die im Dunkeln durch einen Second Messenger (cGMP) dazu angeregt werden ständig offen zu bleiben. Dadurch strömen positiv geladene Ionen in die Zellen ein. Gleichzeitig besitzen Photorezptoren einen Pumpmechanismus durch den Natrium-Ionen beständig aus der Zelle hinaustransportiert werden. Im Dunkeln stellt sich ein Gleichgewicht zwischen Na+Einstrom und Na+ Ausstorm ein, so dass die Membran des Photorezeptors relativ hoch ist (nur ca -20 bis -30 mV). Diese Bewegung positiver Ladungen durch die Membran, die in der Dunkelheit stattfindet, wird als Dunkelstrom bezeichnet. Im Dunkeln setzt der Photorezeptor an seiner synaptischen Endigung beständig den Neurotransmitter Glutamat frei. Wenn wir nachts im dunklen Zimmer schlafen, kann das Stunden andauern. Die Photorezeptorsynapse ist deshalb darauf spezialisiert, einen grossen Nachschub an Glutamat Transmittern bereitzustellen.
Wenn Licht auf die Photorezeptoren fällt, dann wird es von den lichtempfindlichen Photopigmenten in den Membranscheibchen absorbiert (bei Stäbchen: Rodopsin). Durch die Absorption wird eine Signaltransduktionskaskade in Gang gesetzt, die zur Aktivierung eines Enzyms führt, das den Second Messenger cGMP abbaut. Wenn die Konzentration von cGMP fällt, dann schliessen die Na+ Kanäle. Dadurch wird der Na+ Einstrom unterbrochen, wohingegen der Na+ Ausstrom über Pump-Mechanismen weiterhin bestehen bleibt. Bei einem Lichtreiz ist also der Na+ Ausstrom gösser als der Einstrom, wodurch das Membranpotential negativer wird, d.h. der Photorezeptor wird hyperpolarisiert. Je stärker der Lichtreiz, desto stärker ist die Hyperpolarisation. Durch die Hyperpolarisation wird die Neurotransmitter-Ausschüttung an der Synapse zunehmend vermindert, so dass kein Glutamat mehr freigesetzt wird.
Unsere Photorezeptoren sind relativ träge. Sie brauchen einige Zeit, um auf einen Lichtreiz zu reagieren. Fällt ein zweiter Lichtreiz auf einen Photorezeptor, bevor die Antwort auf den ersten Lichtreiz beendet ist, verschmelzen die beiden Antworten. Wir können dann die beiden Reize nicht mehr getrennt wahrnehmen. Die Trägheit unserer Photorezeptoren bildet die Grundlage unserer Film- und Fernsehtechnik. Folgen die Bilder genügend schnell aufeinander (etwa 50 Bilder pro Sekunde), entsteht für uns der Eindruck einer fliessenden Bewegung ohne Flimmern.
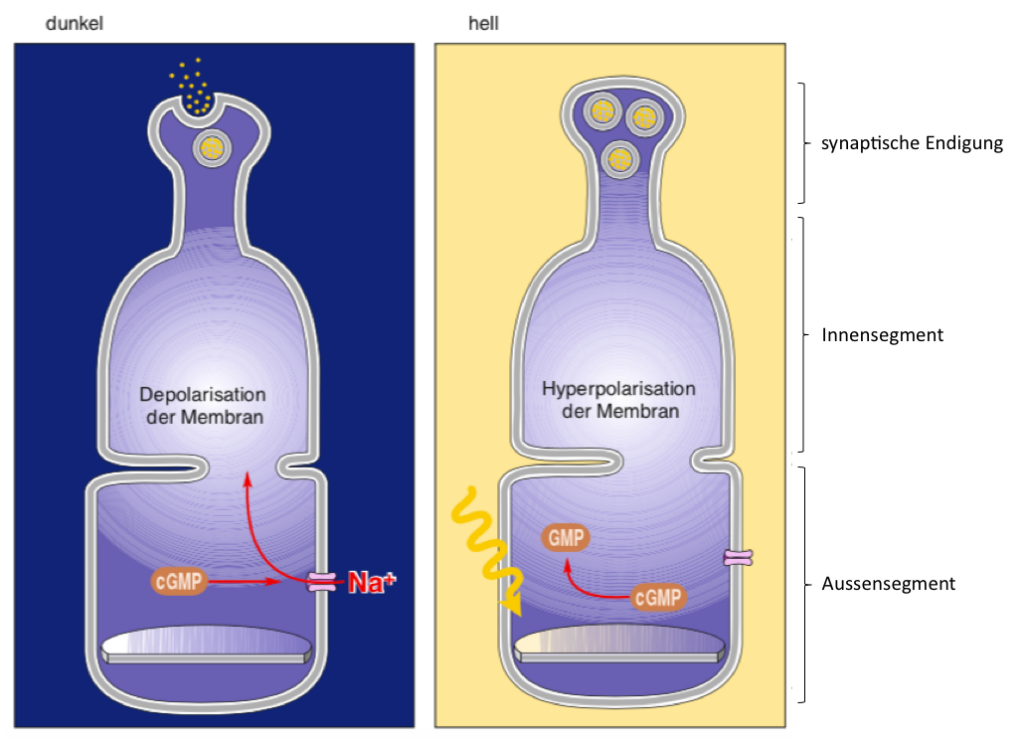
Abbildung 9.9 – Hyperpolarisierende Lichtreaktion der Photorezeptoren. Photorezeptoren sind in der Dunkelheit aufgrund des Natriumeinstroms, des sogenannten Dunkelstroms, dauerhaft depolarisiert. Licht führt zur hyperpolatisation der Zelle. Quelle: Mark Bear et al.: Neurowissenschaften, 4.Auflage, Springer Spektrum, 2018, S. 330
9.7.1 – Neuronale Verschaltung und laterale Hemmung
Photorezeptoren bilden Synapsen mit nachgeschalteten Zellen. Ein wichtiger Zelltyp sind Bipolarzellen, die über die Synapse Information von den Photorezeptoren erhalten. Der Photorezeptor schüttet im Dunkeln sehr viel Glutamat als Neurotransmitter aus, welches dann die Dendriten der Bipolarzellen erreicht. Man unterscheidet zwei Typen der Bipolarzellen: ON-Bipolarzellen haben spezielle dendritische Rezeptoren, so dass die Bipolarzelle durch den Glutamat-Neurotransmitter gehemmt wird. Fällt ein Lichtstimulus auf den Photorezeptor, dann wird die Glutamat-Auschüttung vermindert, damit fällt die Hemmung der ON-Bipolarzelle weg und es kommt zu einer Depolarisation. Das heisst, die ON-Bipolarzelle wird depolarisert, wenn ein Lichtstrahl auf den vorgeschalteten Photorezeptor fällt – die ON-Bipolarzelle wird durch den Lichtreiz quasi „angeschaltet“.
OFF-Bipolarzellen zeigen ein genau gegensätzliches Verhalten. Wenn der Glutamat-Neurotransmitter auf die dendritischen Rezeptoren der OFF-Bipolarzellen trifft, wird die Bipolarzelle erregt. Wenn jetzt ein Lichtstrahl auf den Photorezeptor trifft, wird die Glutamatauschüttung vermindert, dadurch wird die Erregung der OFF-Bipolarzelle reduziert und es kommt zur Hyperpolarisation. Das heisst, die OFF-Bipolarzelle wird hyperpolarisert, wenn ein Lichtstrahl auf den vorgeschalteten Photorezeptor fällt – die OFF-Bipolarzelle wird durch den Lichtreiz quasi „ausgeschaltet“.
Bipolarzellen leiten ihre Information über eine erregende Synapse an nachgeschaltete Ganglienzellen weiter. Die Ganglienzellen sind sozusagen die „Endabnehmer“ der in der Netzhaut verarbeiteten Information und generieren Aktionspotentiale, die dann über den Sehnerv das Auge verlassen und zum Gehirn weitergeleitet werden.
Signalverarbeitung in den Zellen der Retina
Im Zeitintervall t1 bis t2 wurde ein Photorezeptor belichtet. Wie reagieren die verschiedenen Zelltypen der Retina auf den Lichtimpuls? Achtung: in dieser Abbildung sind die Photorezeptoren so angeordnet, dass das Aussensegment oben und die synaptische Endigung unten zu sehen ist. Ziehen sie die Bilder der elektrischen Ableitungen an die entsprechenden Zellen. Sie können die Übung oben rechts vergrössern.
Ganglienzellen haben besondere rezeptive Felder, die aus einem Zentrum und einem ringförmigen Umfeld bestehen. Die Ganglienzelle wird vor allem von dem Photorezeptor im Zentrum des rezeptiven Feldes beeinflusst – man spricht hier von ON-zentrum Ganglienzellen und OFF-Zentrum Ganglienzellen. Zusätzlich erhält die Ganglienzelle auch Information von lateral gelegenen Photorezeptoren, aus dem Umfeld ihres rezeptiven Feldes. Dabei verursacht ein Lichtreiz im Umfeldes und ein Lichtreiz im Zentrum immer entgegengesetzte Reaktionen der Ganglienzelle: eine ON-Zentrum Ganglienzelle reagiert auf einen Lichtreiz im Zentrum mit einer erhöhten Aktionspotentialfrequenz (sie wird quasi „angeschaltet“), aber auf einen Lichtreiz im Umfeld mit einer geringen Aktionspotentialfrequenz. Wozu braucht es ein solch kompliziertes Konstrukt mit ON-Zentrum und OFF-Zentrum Ganglienzellen, die jeweils entgegengesetzte Umfelder haben? Ganz einfach, die Zellen sind keine Helligkeitsdetektoren sondern Kontrastdetektoren. Sie erkennen den Kontrast zwischen dem Zentrum und dem Umfeld. Diese Funktion ist wichtig, um Objekte voneinander zu trennen und getrennt erkennbar zu machen, selbst wenn die Objekte sehr ähnlich sind.
Ein eindrücklichen Beispiel für den Mechanismus der Kontrastverschärfung finden Sie in Abbildung 9.10. Dort sehen Sie einen grauen Fleck entweder auf schwarzem oder weissem Hintergrund. Entlang der Hell-Dunkel-Grenze ist der hellere Teil jeweils etwas heller und der dunklere etwas dunkler als die weitere Umgebung. Entsprechend hat man bei einem schwarzen Hintergrund das Gefühl, dass der graue Fleck einen hellen Rand hat, wohingegen man bei dem weissen Hintergrund einen dunklen Rand wahrnimmt. Bestimmt man die Helligkeit der beiden Beispiele mit einem Messgerät und trägt sie in einem Diagramm auf, erhält man lediglich einen Helligkeitsunterschied zwischen dem grauen Fleck und dem Hintergrund, ohne helle oder dunkle Ränder. Sprich: tatsächlich existieren diese Ränder nicht. Aber woher kommen sie dann? Die Ränder sind ein reines Produkt unseres Wahrnehmungsvorgangs.
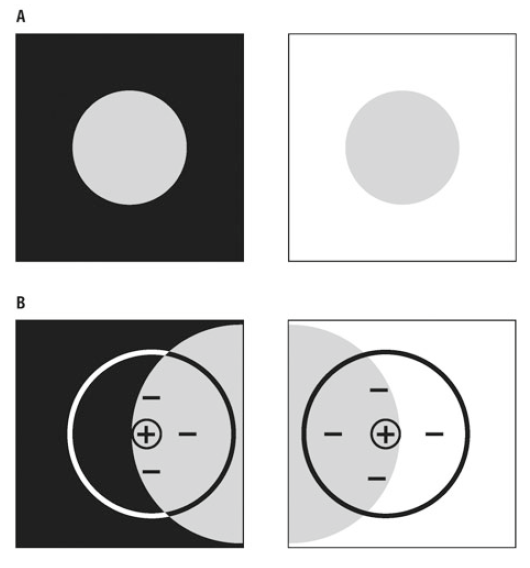
Abbildung 9.10 – Beispiel veränderter Wahrnehmung bei Simultankontrasten und Erklärung durch rezeptive Felder/laterale Hemmung. Aus Physiologie des Menschen, R.F. Schmidt, F. Lang, M. Heckmann, 31. Aufl, 2010, Abb. 18.15
Der untere Teil der Abbildung zeigt ein Beispiel für ein ON-Zentrum rezeptives Feld. ON-Zentrum Zellen werden durch Licht im Zentrum erregt, durch Licht im Umfeld aber gehemmt. Wenn also des Zentrum des rezeptiven Feldes auf den grauen Fleck gerichtet ist, wird die ON-Zentrum Zelle entsprechend erregt. Ist das Umfeld sehr dunkel (schwarzer Hintergrund) ist die Hemmung der ON-Zentrum Zelle so gering, dass der Rand des grauen Flecks heller erscheint. Ist das Umfeld hingegen sehr hell (weisser Hintergrund), dann wird die Aktivität der ON-Zentrum Zelle stark gehemmt und man nimmt einen dunklen Rand wahr.
An diesem Beispiel sehen Sie, dass das Gehirn sehr empfindlich auf Kontraste reagiert. Wenn Zentrum und Umfeld unterschiedlich hell sind, reagiert die Zelle; und zwar umso heftiger, je stärker der Unterschied ist. Ihr Sehsystem sagt Ihnen, was Sie wissen müssen: Hier ist die Trennlinie zwischen zwei grauen Flächen, die sich nur wenig unterscheiden. Als ob es um jedes Objekt herum mit einem Stift eine Umrandung zeichnen würde und Ihnen so hilft, die Objekte zu trennen.
9.8 – Ausserhalb des Auges
Axone der Ganglienzellen werden im Sehnerv gebündelt. Hier verlassen die Neuronen das Auge und die visuellen Informationen werden an verschiedene Gehirnregionen mit unterschiedlichen Funktionen weitergeleitet. Manche Projektionsgebiete des Sehnervs sind an der Regulation von biologischen Rhythmen beteiligt, die mit dem Wechsel von Licht und Dunkelheit im Tageszyklus synchronisiert sind. Andere sind an der Kontrolle der Augenposition und der optischen Abbildung im Auge beteiligt. Die Information, die zur bewussten Sehwahrnehmung führt, wird über den Thalamus zum primären visuellen Cortex geleitet, wo sie ausgewertet und gespeichert wird.