Die Konzepte Lernen und Gedächtnis sind die Grundlage für die Anpassung unseres Verhaltens an die Umwelt. Das Aufnehmen, Speichern und Abrufen von Information hilft uns nicht nur dabei Dinge zu erkennen, sondern führt auch dazu, dass wir ein Identitätsgefühl entwickeln.
Lernziele
- Sie können dass Gedächtnis sowohl nach Speicherzeit als auch nach Art der Information kategorisieren.
- Sie können den Informationsfluss zwischen unterschiedlichen Gedächtnisspeichern beschreiben.
- Sie kennen die Rolle des Hippocampus für das deklarative Gedächtnis.
- Sie kennen die Hebb’sche Regel und können sie am Beispiel des klassischen Konditionierens beschreiben.
- Sie können die vermuteten physiologischen Grundlagen des Lernens und Gedächtnisses (Stichwort synaptische Plastizität/LTP) beschreiben.
13.1 – Gedächtniskategorien
Das Gedächtnis kann auf unterschiedliche Art und Weise kategorisiert werden. Zum einen kann man das Gedächtnis inhaltlich bzw. nach der Art der Information kategorisieren, die gespeichert wird. Zum anderen kann man Gedächtnis nach Speicherzeit, das heisst entsprechend der Zeit über die die Gedächtnisprozesse effektiv sind, unterteilen.
13.1.1 – Zeitliche Kategorisierung
Obwohl die Details immer noch Gegenstand aktueller Forschung sind, ist man sich generell einig darüber, dass es drei zeitliche Hauptspeichersysteme gibt. Das sensorische Gedächtnis (manchmal auch als Ultrakurzzeitgedächtnis bezeichnet), das Kurzzeit– bzw. Arbeitsgedächtnis, sowie das Langzeitgedächtnis.
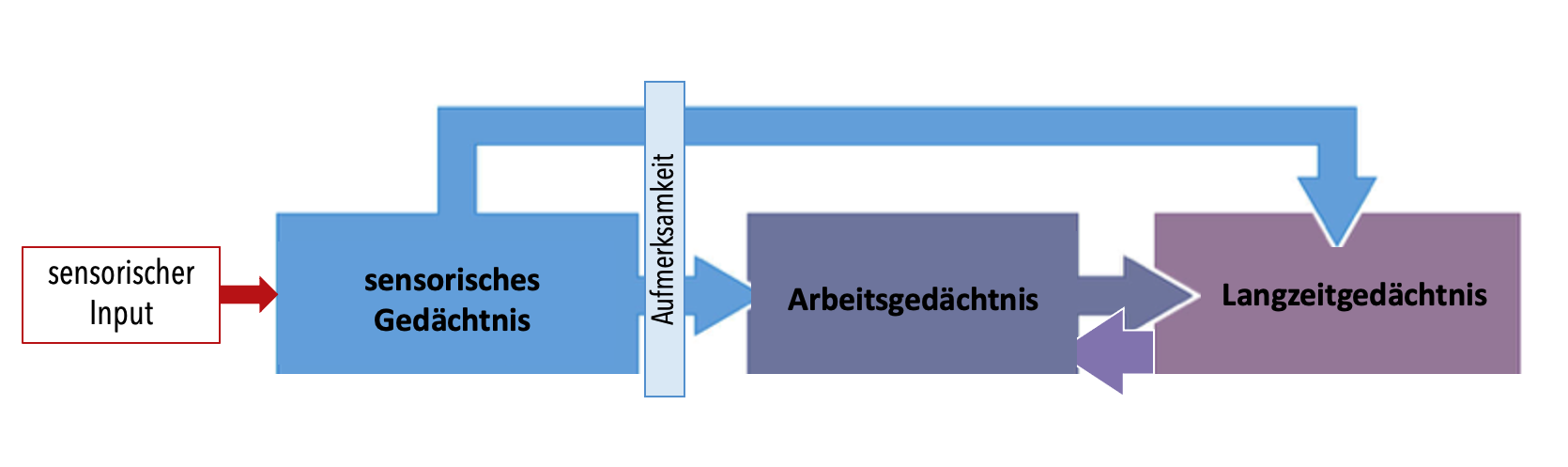
Abbildung 13.1 – Die drei zeitlichen Hauptspeichersysteme des menschlichen Gedächtnisses. Quelle: Dale Purves, Neuroscience, 3. Auflage, Sinauer Associates Inc., 2004, S. 734, verändert.
Reize aus der Umwelt gelangen zunächst ins sensorische Gedächtnis. Der Speicher ist enorm gross, allerdings zeitlich auf wenige hundert Millisekunden begrenzt. Manche der im sensorischen Gedächtnis gespeicherten Informationen gelangen dann – etwa durch Aufmerksamkeitsausrichtung – ins Arbeitsgedächtnis. Der Prozess der Enkodierung beschreibt den Mechanismus, der zur Aufnahme von Information, zu einer dauerhaften Repräsentation und späteren Wiedergabe führt. Da der Organismus kontinuierlich Informationen verarbeitet, ist die mittlere Aufrechterhaltung von Informationen im Arbeitsgedächtnis kurz (einige Sekunden-Minuten). Alltägliche Beispiele für das Arbeitsgedächtnis sind das Behalten einer Telefonnummer, die uns jemand mitteilt, bis wir sie eingetippt haben oder aber eine Einkaufsliste, die man im Kopf erstellt. Experimentell kann das Arbeitsgedächtnis untersucht werden, in dem Personen eine Zahlenliste vorgelesen wird und bestimmt wird, wie viele Zahlen sie wiedergeben können (= Gedächtnisspanne). Für gewöhnlich beträgt die Gedächtnisspanne 7 +/- 2 Informationseinheiten (z.B. Satzteile, Nummerngruppen). Ein wichtiger Aspekt des Arbeitsgedächtnisses ist, dass Informationen nicht nur aufrechterhalten, sondern auch manipuliert werden können (wenn uns beispielsweise eine Zahlenreihe genannt wird, können wir diese «im Kopf» umdrehen und rückwärts wiedergeben).
⇒Ziehen Sie den Regler von links nach rechts. Werfen Sie zunächst auf eine Reihe einen einmaligen Blick während Sie den Regler von links nach rechts schieben und versuchen Sie direkt danach die Zahlenfolge aufzuschreiben. Dann wiederholen Sie diesen Prozess für die nächste Reihe! Wie viele Zahlen können Sie mit einmaligem Schieben pro Reihe notieren? Schaffen Sie 7+/-2 Zahlen?
Was für eine Strategie haben Sie angewendet, um möglichst viele Zahlen wiedergeben zu können? Haben Sie versucht, die Information aktiv aufrecht zu erhalten durch kontinuierliche Auffrischung/Wiederholung der Zahlen?
Ein Teil der Information wird schliesslich aus dem Arbeits- ins Langzeitgedächtnis, ein dauerhaftes Speichersystem, übertragen. Das kann zum Beispiel durch Üben (also sehr aufmerksames Wiederholen, wird auch manchmal «rehearsal» genannt) und damit korrespondierendes Zirkulieren der Information erreicht werden. Das Überführen von einer kurzfristigen in eine weniger störbare und langfristigere Gedächtnisspur nennt man Konsolidierung. Die im Langzeitgedächtnis geformte Gedächtnisspur nennt man Engramm. Eine solche Gedächtnisspur ist nicht fix und stabil, sondern das Gedächtnis ist ein dynamischer Prozess, der sich immer wieder verändert – so kann durch jede Wiedergabe oder Teilwiedergabe eines Gedächtnisinhaltes, die Gedächtnisspur verändert (z.B. mit neuen Informationen ergänzt werden) und gestärkt werden. Diesen Prozess bezeichnet man als Rekonsolidierung. Wenn wir Informationen aus dem Langzeitgedächtnis abrufen und wiedergeben, wird die Information aus dem Langzeitspeicher wieder ins Arbeitsgedächtnis gebracht, wo sie bis zur Wiedergabe auch ohne Wiederholung «am Leben» erhalten werden kann. Anders als für das sensorische und für das Arbeitsgedächtnis ist eine Abschätzung der genauen Speicherzeit und Speicherkapazität des Langzeitgedächtnisses schwierig und immer noch Gegenstand aktueller Forschung.
13.1.2 – Kategorisierung nach Art der Information
Ganz grundsätzlich kann man zwei qualitativ unterschiedliche Langzeitgedächtnisspeicher unterscheiden – das deklarative (auch explizites oder Wissens-Gedächtnis genannt) und das non-deklarative Gedächtnis (auch implizites oder Verhaltens-Gedächtnis genannt).
Deklaratives Gedächtnis
Auf deskriptiver Ebene umfasst das deklarative Gedächtnis verbalisierbares Wissen über Fakten und Ereignisse. Wir können dieses Wissen bewusst und intentional abrufen und wiedergeben. Das deklarative Gedächtnis wurde klassisch eingeteilt in das episodische Gedächtnis (manchmal auch autobiographisches Gedächtnis genannt) und das semantische Gedächtnis. Das episodische Gedächtnis beinhaltet Erinnerungen an Ereignisse und deren räumlich-zeitliche Einbettung. Wir können uns zum Beispiel an eine Wanderung auf den Säntis (wo) im Sommer vor zwei Jahren (wann) erinnern. Das semantische Gedächtnis bezieht sich auf Sachwissen und Fakten ohne räumlich-zeitliche Einbettung. Wir wissen beispielsweise, dass der Säntis ein Berg in der Schweiz ist, wissen aber vermutlich nicht mehr, wann und wo wir das genau gelernt haben. Ausserdem beinhaltet das semantische Gedächtnis die Fähigkeit Familie, Freunde oder Bekannte zu erkennen. Heute ist die strikte Trennung zwischen episodischem und semantischem Gedächtnis eher eine einleuchtende deskriptive und weniger funktionale Unterscheidung von zwei Formen eines Gedächtnisses.
Non-deklaratives Gedächtnis
Das non-deklarative Gedächtnis ist eine unbewusste, nicht intentionale Gedächtnisform. Als non-deklarativ wird sie deswegen bezeichnet, weil Inhalte nicht verbalisierbar sind, das heisst nicht berichtet werden können. Motorische und kognitive Fertigkeiten (= prozedurales Gedächtnis), wie die Fähigkeit Sprache zu gebrauchen oder aber Fahrrad zu fahren sind zwei Beispiele dieser Gedächtnisform. Eine weitere Unterkategorie ist das Priming («Bahnung»). Priming meint die schnellere und/oder genauere Verarbeitung als Konsequenz einer vorangegangenen Verarbeitung eines Ereignisses. Auch einfaches assoziatives Lernen (wie etwa die klassische Konditionierung) und nicht-assoziatives Lernen (Habituation und Sensitivierung) wird dem non-deklarativen Gedächtnis zugeordnet. Während Habituation bedeutet, dass wir weniger responsiv auf eine wiederholte Darbietung eines als eher unwichtig erkannten Reizes reagieren (wir spüren beispielsweise direkt nach dem Anziehen die Kleidung auf unserer Haut, wir nehmen diese Empfindung aber relativ schnell nicht mehr oder weniger stark war), bedeutet eine Sensitivierung, dass wir stärker auf eine wiederholte Darbietung eines besonders intensiven oder noxischen Reizes reagieren (ein Beispiel ist die Sensitivierung von Schmerzempfinden).
Ein wichtiger Unterschied zwischen dem deklarativen und non-deklarativen Gedächtnis ist, dass die Inhalte in unterschiedlichen Hirnarealen und neuronalen Strukturen beherbergt werden (siehe nachfolgende Abbildung). Für die Bildung neuer Gedächtnisinhalte des deklarativen Gedächtnisses scheint vor allem der Hippocampus eine wichtige Rolle zu spielen.
13.2 – Der Hippocampus und das deklarative Gedächtnis
13.2.1 – Der Fall H.M.
Wie schon im Kapitel Aufmerksamkeit beschrieben, liefern häufig Patienten mit gewissen Hirnläsionen wichtige Erkenntnisse über höhere Hirnfunktionen, so auch über das Gedächtnis. Der Patient H.M. (1926-2008) war einer der Ausgangspunkte für eine systematische Gedächtniskategorisierung auf neurobiologischer Basis und zeigte, wie wichtig der Hippocampus (sowie umliegende Areal im medialen Temporalcortex) für bestimmte Gedächtniskategorien ist. H.M. litt seit seiner Kindheit an Epilepsie. Da seine Erkrankung medikamentös nicht unter Kontrolle gebracht werden konnte, wurde ihm 1953 beidseitig der mediale Temporalcortex (inklusive grosser Teile des Hippocampus und parahippocampalen Cortex sowie der Amygdala, Abbildung 13.2, links) entfernt. Diese OP führte zu einem unerwarteten Ergebnis: H.M. konnte sich fortan keine neuen Informationen langfristig aneignen und wiedergeben, er litt an einer anterograden Amnesie (= Störung der Neubildung des deklarativen Gedächtnisses, Abbildung 13.2, rechts). Beispielhaft für das Ausmass seiner Amnesie war, dass er auch nach 6 Jahren unmittelbar nach seiner Hirnoperation eingezogene Hausnachbarn nicht erkannte, sich immer wieder am gleichen Puzzle erfreuen konnte und dass er schon kurze Zeit nach Beendigung der Mittagsmahlzeit nicht mehr wusste, ob er schon zu Mittag gegessen hatte oder nicht. Allerdings zeigte er keine Intelligenz- oder Persönlichkeitsveränderungen, auch keine Veränderungen in der Motivation, Wahrnehmung oder im abstrakten Denken. Obwohl auch seine Erinnerungen an Geschehnisse aus der kürzeren Vergangenheit teilweise beeinträchtigt waren, waren seine Erinnerungen an Jahre zurückliegende Ereignisse lebendig und offensichtlich ohne Lücken. Er konnte ausserdem auch neue Fertigkeiten erlernen, was zeigt, dass sein non-deklaratives Gedächtnis intakt war.
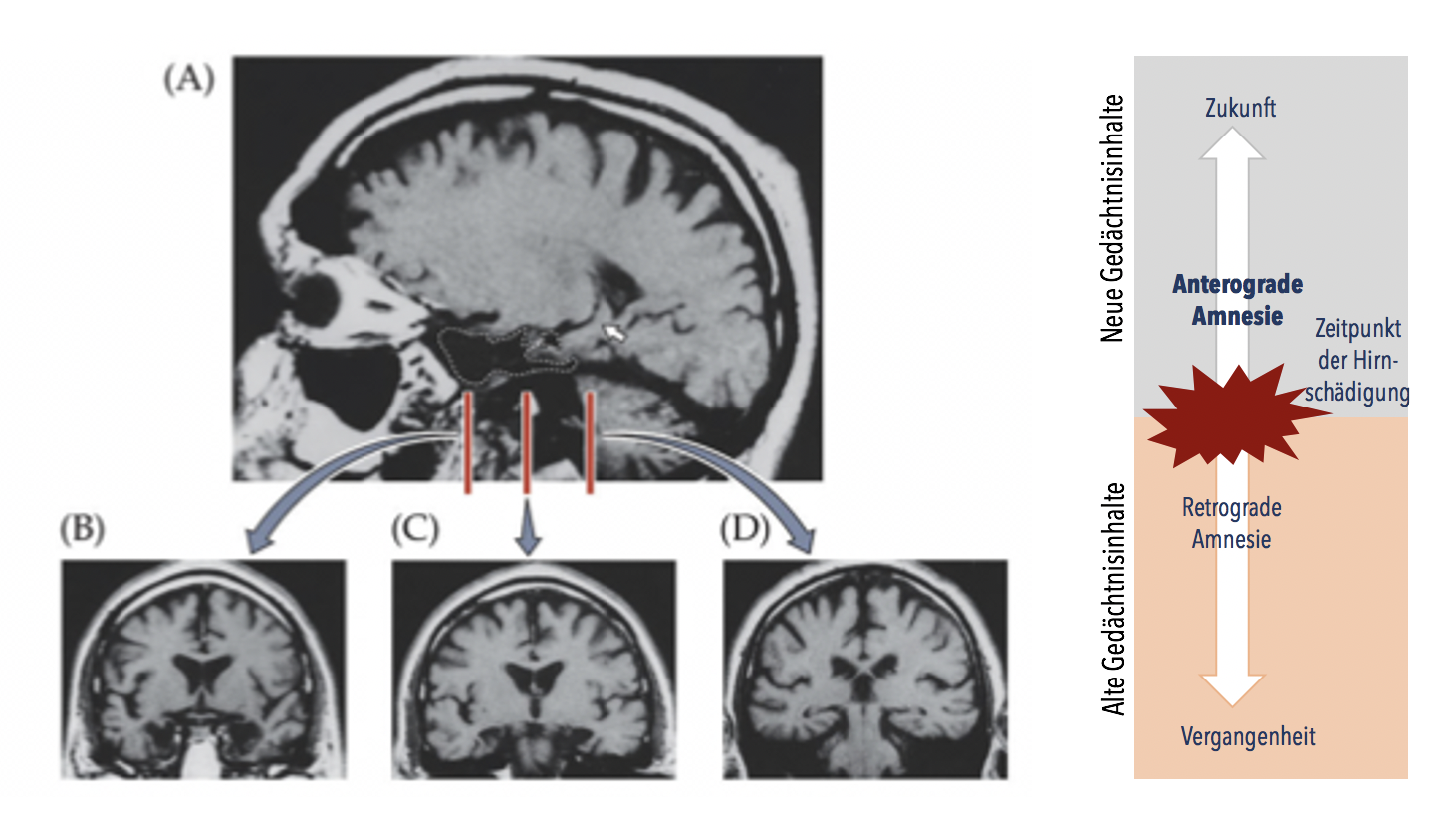
Abbildung 13.2 – Links: MRT-Bilder von H.M.s Gehirn nach der Entfernung des Hippocampus und der umliegenden Areale. Quelle: Dale Purves, Neuroscience, 3. Auflage, Sinauer Associates Inc., 2004, S. 743. Rechts: Mögliche Konsequenzen einer Hirnläsion auf das Erinnern alter und das Bilden neuer Gedächtnisinhalte. Schädigungen des Hippocampus und der umliegenden Areale können zu einer anterograden Amnesie führen. Quelle: Eigene Darstellung 2020.
13.3 – Assoziatives Lernen
Lernen und Gedächtnis sind eng miteinander verknüpft. Ein allgemeiner Mechanismus, der dem Lernen zugrunde liegt, ist die Bildung von Assoziationen. Bei einer Assoziation handelt es sich um eine enge zeitliche Paarung von Information bzw. Reizen. Obwohl Assoziationen im weiteren Sinne generell für den Wissenserwerb (also auch für das deklarative oder Wissensgedächtnis) wichtig sind, besteht eine Assoziation im engeren Sinne beim assoziativen Lernen in der Herstellung einer Verbindung zwischen Reizen (S) und Reaktionen (R). Zwei Beispiele für assoziatives Lernen sind die klassische Konditionierung und die operante Konditionierung.
13.3.1 – Klassische Konditionierung
Bei der klassischen Konditionierung basiert eine Assoziation darauf, dass die auftretenden Reize zeitlich simultan oder kurz aufeinander folgen (= Kontiguitätsprinzip). Bei dieser Form des Lernens wird ein unkonditionierter Reiz, der eine unkonditionierte Reaktion hervorruft (eine sich natürliche ereignende, meist angeborene Reaktion) wiederholt mit einem neutralen Reiz, der diese Reaktion nicht hervorruft, kombiniert. Diese Lernform geht auf den Physiologen Iwan Pawlow und sein bekanntestes Beispiel, das des Pawlowschen Hundes, zurück. Pawlow vermutete, dass bereits die Schritte des Besitzers eines Zwingerhundes ausreichen, um einen Speichelfluss beim Hund auszulösen, da dem Geräusch der Schritte regelmässig die Fütterung folgte. Um dies experimentell zu zeigen, führte er folgendes Experiment durch: Zunächst bot er einem Hund Nahrung dar (unkonditionierter Reiz, US) was natürlicherweise Speichelfluss auslöst (UR, unkonditionierte Reaktion). Dann hat Pawlow das Futter, den US, der den Speichelfluss (UR) auslöst, wiederholt zusammen mit einem neutralen Reiz (NS, hat keine Reaktion zur Folge), einer Glocke, dargeboten. Durch diese wiederholte, zeitgleiche Darbietung wurde die Glocke zu einem konditionierten Reiz (CS), der nun auch alleine den Speichelfluss (konditionierte Reaktion, CR) auslösen konnte (Abbildung 13.4; auch das Futter (US) löst natürlich nach dieser Konditionierungsphase noch immer den Speichelfluss aus (UR)). Entscheidend für die Stärke der Assoziation ist 1. die Häufigkeit der gemeinsamen Darbietung des ursprünglich neutralen Reizes (NS, der dann zu CS wird) und des unkonditionierten Reizes (US, der die unkonditionierte Reaktion auslöst), und 2. die zeitliche Kontiguität zwischen dem konditionierten (CS) und dem unkonditioniertem Reiz (US). Optimal sind 500ms. Heute weiss man ausserdem, dass der konditionierte Reiz (CS) eine zuverlässige Vorhersage für das Auftreten des unkonditionierten Reizes (US) erlauben muss, damit es zur Konditionierung kommt.
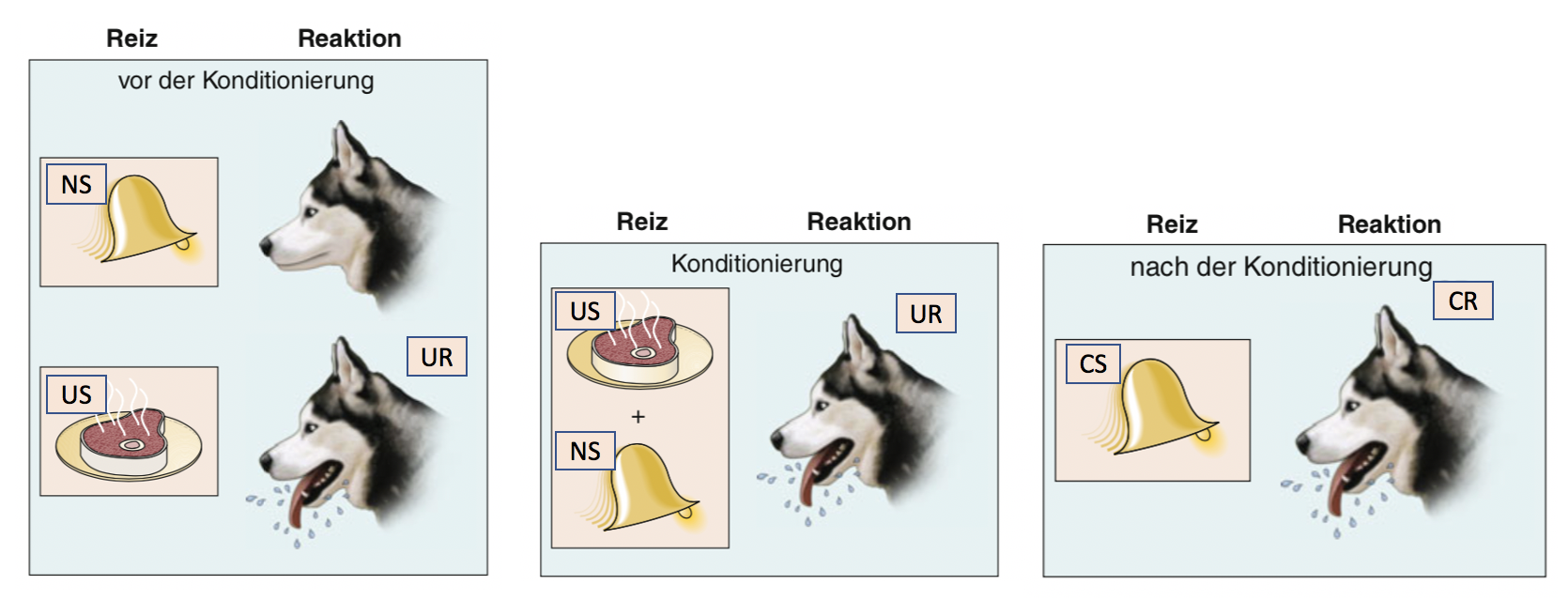
Abbildung 13.3 – Klassische Konditionierung. Quelle: Michael Baer et al., Neurowissenschaften, 4.Auflage, Springer Spektrum, 2018, S. 898.
13.3.2 – Operante Konditionierung
Beim klassischen Konditionieren wird die unkonditionierte Reaktion quasi passiv erlernt. Bei der operanten Konditionierung wird neues Verhalten aktiv erworben. Die operante Konditionierung geht auf Experimente von Burrhus Skinner (Stichwort Skinner-Box) und Edward Thorndike zurück und basiert auf dem Kontingenzprinzip, d.h. einer Verbindung aus einem auslösendem Reiz (S), einer Reaktion (R), und der davon ausgelösten Konsequenz (K): In einer bestimmten Situation folgt unmittelbar auf ein Verhalten eine belohnende oder bestrafende Konsequenz, die zu einer Verstärkung oder Unterdrückung des Verhaltens führt. Das heisst, die Konsequenzen, die auf ein bestimmtes Verhalten folgen, bestimmen die Wahrscheinlichkeit, mit der ein bestimmtes Verhalten unter bestimmten Bedingungen wiederholt werden wird. Das Verhalten, das eine gewisse Konsequenz nach sich zieht, wird häufig zunächst unbeabsichtigt oder rein zufällig gezeigt.
13.3.3 – Extinktion
Wenn im Falle der klassischen Konditionierung der konditionierte Reiz (CS) wiederholt ohne den unkonditionierten Reiz (US) dargeboten wird, wird die konditionierte Reaktion allmählich schwächer oder erlöscht gänzlich. Wenn ähnlich hierzu bei der operanten Konditionierung auf das Verhalten überhaupt keine Konsequenz mehr erfolgt, wird auch diese konditionierte Reaktion allmählich abgeschwächt oder verschwindet (= Prozess der Extinktion).
13.4 – Physiologische Grundlagen: Die Plastizität des Gehirns
Unser zentrales Nervensystem besteht aus einer Vielzahl an Neuronen und noch viel mehr Synapsen, die diese Neurone miteinander verbinden und die über die Informationsübertragung entscheiden. Lernprozesse sind Ausdruck der Plastizität unseres Nervensystems. Das bedeutet, dass der Erwerb eines neuen Verhaltens in der Regel mit der Veränderung von Neuronen und Synapsen einhergeht.
13.4.1 – Hebb’sche Regel
Die Hebb’sche Regel stellt eine neurophysiologische Grundlage der Bildung von Assoziationen dar und besagt Folgendes: «Wenn ein Axon des Neurons A nahe genug an einem Neuron B liegt, so dass Zelle B wiederholt oder anhaltend von Neuron A erregt wird, so wird die Effizienz von Neuron A für die Erregung von Neuron B durch einen Wachstumsprozess oder eine Stoffwechseländerung in beiden oder einem der beiden Neurone erhöht». Das heisst, dass die Verbindung zwischen Neuronen, die zeitgleich aktiv sind und feuern, stärker wird («Neurons that fire together wire together»).
Wenn wir die klassische Konditionierung des Pawlowschen Hundes als Beispiel nehmen, kann man sich diesen Prozess folgendermassen vorstellen: Generell sind zwei präsynaptische (Synapse 1 und 2) und ein postsynaptisches Neuron beteiligt. Synapse 1 wird durch die Glocke, einen neutralen Reiz erregt. Dieser Ton reicht nicht aus, um das postsynaptische Neuron, an der Synapse 1 und 2 konvergieren, zum Feuern zu bringen. Nun wird Synapse 2 durch die Darbietung des Futters (unkonditionierter Reiz), erregt, das im postsynaptischen Neuron die Aktivierung des Speichelflusses (unkonditionierte Reaktion) auslöst. Wenn nun Glocke und Futter gemeinsam dargeboten werden, das heisst Synapse 2 gleichzeitig oder kurz nach Synapse 1 erregt wird, verstärkt das Feuern des postsynaptischen Neurons, ausgelöst durch Synapse 2, die Aktivität aller Synapsen, die an diesem postsynaptischen Neuron gleichzeitig aktiv waren – das heisst auch die Erregbarkeit von Synapse 1. Nach mehreren Paarungen von Glocke und Futter wird Synapse 1 zunehmend stärker und die Darbietung der Glocke (konditionierter Reiz) allein reicht aus, um das postsynaptische Neuron zum Feuern zu bringen und damit den Speichelfluss auszulösen (konditionierte Reaktion). ACHTUNG: das ist ein stark vereinfachtes, konzeptuelles Beispiel. Tatsächlich laufen bei der Konditionierung kompliziertere Prozesse ab.
⇒Das Video erklärt dir an einem stark vereinfachten konzeptuellen Beispiel der klassischen Konditionierung das Prinzip der Hebb-Regel.
Quelle: Eigene Kreation, 2020
13.4.2 – Langzeitpotenzierung (LTP): eine Form der synaptischen Plastizität
3 LTP-Eigenschaften
Die LTP hat 3 wichtige Eigenschaften, die sie zu einem attraktiven neuronalen Mechanismus für Informationsspeicherung macht:
- Zustandsabhängigkeit: Der Zustand des Membranpotentials des postsynpatischen Neurons bestimmt, ob LTP auftritt oder nicht. Eine einzelne Stimulation des präsynatpischen Neurons würde normalerweise keine LTP auslösen. Wenn diese Stimulation allerdings mit einer starken Depolarisation des postsynaptischen Neurons gepaart auftritt, kommt es zu LTP. Allerdings nur dann, wenn die starke Depolarisation innerhalb von 100 ms der präsynaptischen Aktivität stattfindet (Hebb: Neurons that fire together, wire together).
- Inputspezifität: Wenn die LTP durch die Stimulation einer Synapse induziert wird, tritt sie nicht in anderen, inaktiven Synapsen desselben Neurons auf. Dies könnte wichtig sein für eine Speicherung spezifischer Information.
- Assoziativität: eine schwache Stimulation einer Synapse allein triggert keine LTP. Wenn allerdings eine benachbarte Synapse zur gleichen Zeit stark stimuliert wird, kommt es zur LTP in beiden synaptischen Pfaden. Das heisst, ein Set an Information könnte mit einem anderem Set an Informationen verbunden werden (siehe konzeptuelles Beispiel zur Konditionierung).
Molekulare Mechanismen der LTP
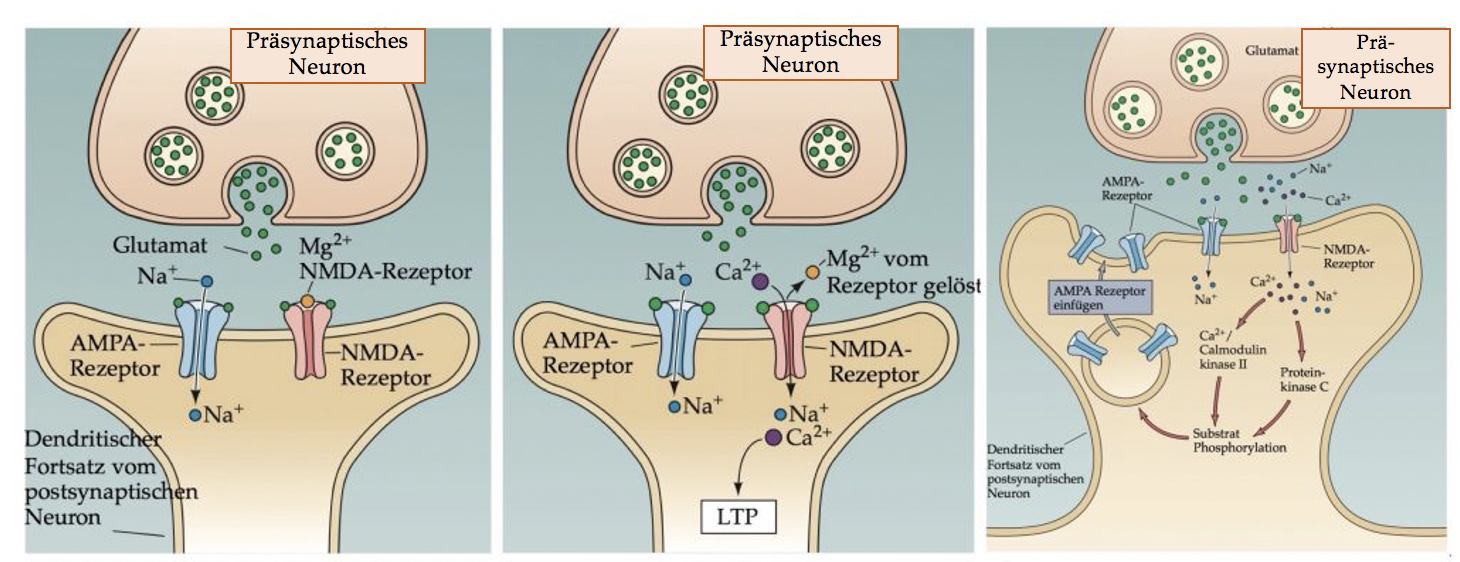
Abbildung 13.4 – Darstellung der synaptischen Vorgänge, die der LTP zugrunde liegen. Links: normales Ruhepotential, Mitte: während postsynaptischer Depolarisation; Rechts: Bildung neuer AMPA-Rezeptoren – ein LTP-Mechanismus. Quelle: Dale Purves, Neuroscience, 3. Auflage, Sinauer Associates Inc., 2004, S. 588. Verändert 2020.
An den Dendriten der Pyramidenzellen im Hippocampus gibt es Glutamatrezeptoren vom NMDA- und vom AMPA-Typ (Abbildung 13.5). Ein einzelnes Aktionspotenzial verursacht die Ausschüttung einer Glutamatmenge, die zur Öffnung einiger Kanäle vom AMPA-Typ ausreicht. Die NMDA-Kanäle öffnen bei normalem Ruhepotential des postsynaptischen Neurons nicht, weil sie durch Magnesium-Ionen (Mg2+) blockiert sind; da sich nur die AMPA-Kanäle öffnen, ergibt sich ein relativ kleines exzitatorisches postsynaptisches Potential (EPSP). Da die NMDA-Kanal-Blockade durch Mg2+ aber spannungsabhängig ist, ändert sich die Funktion der Synapse, wenn die postsynaptische Zelle depolarisiert ist. Kommt es zum Beispiel zu einer Depolarisation der postsynaptischen Membran durch wiederholte, hochfrequente Stimulation (Serie von Aktionspotentialen) oder zur Depolarisation, wenn mehrere konvergierende Synapsen gleichzeitig aktiv sind, dann wird das Mg2+ Ion vom NMDA-Rezeptoren abgestossen und der NMDA-Kanal ist geöffnet. Dadurch kann Ca2+ ins postsynaptische Neuron einströmen, was zu einer erhöhten Ca2+ Konzentration führt. Dies wiederum aktiviert verschiedene Enzymsysteme und triggert die LTP. Beispielsweise werden neue AMPA-Rezeptoren durch diese Enzymprozesse in den dendritischen Dornfortsatz eingefügt und dadurch die Glutamatsensitivität erhöht (Abbildung 13.5, rechts). NMDA-Rezeptoren verhalten sich also wie ein molekulares und-Gate: Der NMDA-Kanal öffnet nur (und induziert LTP), wenn Glutamat an NMDA Rezeptoren gebunden werden kann und die postsynaptische Zelle depolarisiert ist (keine Mg2+-Blockade der Kanäle). Der NMDA-Rezeptor detektiert also das gleichzeitige Feuern zweier Neurone, die durch ein Synapse verbundene sind – er fungiert als Koinzidenzdetektor.
Um synaptische Stärkung nützlich zu machen, müssen andere Prozesse selektiv spezifische Synapsen schwächen. Die Langzeitdepression (LTD) ist ein solcher Prozess.
13.4.3 – Langzeitdepression (LTD): der Gegenspieler der LTP
Unter LTD versteht man eine lang andauernde Depression bzw. Abschwächung der Signalübertragung an einer Synapse. Es ist wichtig, dass überflüssige synaptische Verbindungen, die für ein gewünschtes Verhalten nicht gebraucht werden, abgebaut werden. An manchen Synapsen benötigt LTP kurze, hochfrequente Stimulation, während LTD auftritt, wenn präsynaptische Axone für längere Zeit (etwa 10-15 min) niederfrequent mit ca. 1 Hz stimuliert werden. Dieses Aktivitätsmuster reduziert postsynaptische Potentiale für mehrere Stunden und ist, ähnlich wie die LTP auch, spezifisch für die aktivierte Synapse.
LTD kann den Anstieg in synaptischer Übertragungsstärke durch LTP auslöschen. Umgekehrt kann auch LTP die Reduktion in synaptischer Übertragungsstärke durch LTD auslöschen. LTP und LTD können somit Synapsen langfristig im Sinne eines Lerneffektes in ihrer Effektivität verstärken oder vermindern.